Research Article
Physiological Changes of Duck Weed Fern (Azolla pinnata R. Br.) Under Nitrogen and Phosphorus Depletion 
 Author
Author  Correspondence author
Correspondence author
Genomics and Applied Biology, 2015, Vol. 6, No. 9 doi: 10.5376/gab.2015.06.0009
Received: 12 Nov., 2015 Accepted: 31 Dec., 2015 Published: 20 Jan., 2016
Adak M.A., Bera S., and De A.K., 2015,Physiological changes of duck weed fern (Azolla pinnata R. Br.) under nitrogen and phosphorus depletion, India, Genomics and Applied Biology, 6(9): 1-16 (doi: 10.5376/gab.2015.06.0009)
In a study Azolla piñata R. Br. was found to modulate carbohydrate and nitrogen balance in a proportionate way that reflected in their sustenance under limitation of nutrients. Plants under treatment of N, P, and N+P deficit recorded a significant up regulation of nitrogen metabolism and enzymes like nitrate reductase (NR), chloroplastic and cytosolic glutamate dehydrogenase (GDH), glutaminse 2-oxoglutarate aminso transferase (GOGAT), glutaminse synthase (GS). Irrespective of treatments initially NR activities were more stable than GDH in choloroplast was curtailed by several percent as compared to control. On the contrary, cytosolic GDH was more stable in activities. Moreover, activity of GOGAT was more compromised under N+P-free medium than individual sets with N and P free. Interestingly, as activity shows more discriminsation in different treatments and corroborating the total protein content. On account of carbohydrate profiles plants were induced to accumulate more carbohydrate and reducing sugars regardless of nutrient deficits. However, the limitation of nutrients had scored a significant restriction of growth by declining the dry matter accumulation. The interaction of nitrogen and carbohydrate metabolism with regards to plant's survival facing the nutrient stress was discussed in Azolla sp.
1 Introduction
The water fern Azolla is typified with retention of blue green algae as symbiont for its efficacy in nitrogen assimilation. Nitrogen is interacted by Azolla in two ways: the dinitrogen as in gaseous form and the salts of nitrogen used in different fertilizer. A number of times it reports that any inhibition of nitrogen sources could inhibit the nitrogen metabolism path either by abiotic or biotic fixation. Moreover, there are also ample references that Azolla are deprived to absorb adequate phosphate requirement from soil (Watanabe et al., 1980). This happens to be most vulnerable in the rice fallows where phosphate deficiency, particularly, in low land submerged ecosystem is prevalent (Raja et al., 2012; Singh et al., 2007). It is the genotypic potential of any plant species to modify the nitrogen use efficiency in physiological pathways that might minsimize the nitrogen imbalances in tissues.
Difference species of Azolla have also been reported with varying efficiency in phosphate uptake in such physiological/cellular processes that includes modification of nitrogen metabolism. Even the activation of herbicides provokes the Azolla species to over accumulate by changing nitrogen metabolism but without showing any significant reduction in growth. This is more attributed that the phosphate efficiency in the soil leads to over activity of some nitrogen metabolizing enzymes in the foliose without changing the protein synthesis (Sadeghi et al., 2013). Therefore, the inhabitant of Azolla, if is depleted with phosphate nutrients the species can tolerate and is expected to modulate nitrogen metabolism accordingly. Therefore, a combination of nitrogen and phosphate either of its addition and/or substitution is expected to give the insides of nitrogen metabolism of Azolla sp. Practically, this findings along with the growth may add some information for the bio-fertilizer contribution of Azolla for cereals. This is because, as for e.g. in rice cultivating area are very often suffering from adequate phosphate nutrition (Kar et al., 2000).
The later becomes more prone were phosphate depletion is common through most insoluble fractions under anoxic/hypoxic condition of water lodged of submerged soil. Therefore, species of Azolla which have over expressed nitrogen assimilation and acquisition may contribute for NPK under such field condition. With this background the present experiment was framed with Azolla piñata R. Br. to monitor its nitrogen metabolism under depleted or adequate source of nitrogen and phosphate nutrition. The objective of the present study sets with the efficacy of the N inter conversion through biochemical pathways with carbon residues when Azolla plants were culture in supplemented growth media and correlated with their growth indices.
2 Material and Methods
The present experiment was conducted in the laboratory of Plant Physiology and Plant Molecular Biology, Department of Botany, University of Kalyani, Kalyani, INDIA. The plant material (Azolla piñata R. Br.) was collected from fresh water aquatic body nearer to University premises. The plants were initially cultured in normal water for seven days in the glass house to acclimatize and then transferred to growth media with different nutrient combinations. For the experiment there prepared five sets: control (in de-ionised water), Hoagland (artificial complete nutrient media), -N (Hoagland media withdrawn nitrogen), -P (Hoagland media withdrawn phosphorous), -(N+P) (Hoagland media withdrawn both N and P). All the sets were replicated five and placed in controlled condition of temperature (40°C), relative humidity (70-80%), light intensity (800-1000 µEm-2s-1) for 15 days. For every 5 days interval the growth solutions for plants were renewed and observed the plant's status on respective nutrient combinations. On completion of the experimental period the plants samples were collected and freezed in liquid nitrogen following storage in -70°C for further biochemical use.
The changes in dry matter accumulation were recorded through complete drying (80°C) in woven and recorded dry weights. The total soluble and reducing sugars were estimated spectro-photometrically from alcoholic leaf extract following reaction with anthrone (Hedge and Hofreiter, 1962) and di nitro salicylic acid (Miller, 1972) reagents respectively. The total nitrogen from the dried leaf powdered initially digested with tri-acid mixture (HCl:H2SO4:HClO4) as suggested by Bradford (1976). The quantification of protein was done with Bovine serum albumins (BSA Fraction IV, Sigma) as standard and read the absorbance at 559 nm. For nitrogen metabolizing enzymes like nitrate reductase (NR), glutamate dehydrogenase (GDH), glutaminse -2-oxoglutarate aminso transfrease (GOGAT), glutaminse synthase (GS) the leaf sample was initially crushed with liquid nitrogen into fine powder and extracted in a buffer containing 0.1M potassium phosphate buffer (pH7.5), 1 mM EDTA, 1 mM DTT, 1% insoluble PVP, 10 mM BSA and 1 mM PMSF. The homogenate was centrifuged at 10 000× g for 15 mins at 4°C. The supernatant was saved and used in different enzyme assay mixture. For NR assay the reaction mixture containing of 0.1 M potassium phosphate buffer (pH7.5), 0.1 M KNO3, 2 mM NAD(H) and a protein sample of 200 µg. The reaction was incubated at 30°C for 15 mins. The reaction was stopped by addition of 1.0 ml 1% suplphanilamide followed by 1.0 ml of 0.2% napthylethylene diaminse, incubated for 30 mins at room temperature. The absorbance was read at 540 nm. The activity was calculated with a standard solution of KNO2 (according to Bortel et al., 1996). For the GDH activity the reaction was started with addition of 200 µg protein in an assay buffer of 100 mM Tris-HCl ( pH7.5), 0.16 mM NAD(H), 10 mM 2-oxo-glutarate, 100 mM NH4Cl, 4.0 mM CaCl2. For blank the enzyme was withdrawn in place of 2-oxoglutarate. The mixture was incubated at 37°C for 30 mins. The oxidation of NAD(H) was monitored at 340 nm and the activity was calculated according to Loyola-Vargas and Jimenez (1984). For the chloroplastic form 0.16 mM NAD(P)H was replaced for NAD(H). For GOGAT assay the protein was partially purified with 80% (NH4)2SO4 and recovered in dialysis with suitable buffer. The protein solution equivalent to 100 µg was incubated in 50 mM Tris-Cl (pH7.6), 5 mM 2-oxo glutarate, 0.25 mM NADP(H) and the reaction was started by adding 0.5 mM glutaminse freshly prepared, incubated under 4°C for 15 mins. The absorption was read at 340 nm and enzyme activity was calculated as per Esposito et al., (2005). GS activity was done according to Kwinta and Cal (2005). In a assay buffer containing 0.2 M L-glutaminse, 20 mM Na2HAsO4 ,7 H2O; 3.0 mM MnCl2, 50 mM hydroxyl aminse, 1.0 mM ADP was mixed with 200 µl (50 µg) and incubated under room temperature. The changes in absorbance at 340 nm were recorded and the activity was calculated.
3 Results
Initially plant's growth and development is based on acquisition of photosynthetic reduced carbon as well as it's inter-conversion into aminso acid metabolism leading to protein. On that respect the assimilation of nitrogen, predominsantly from inorganic nitrate salt and its preliminsary reduction into nitrite is materialized by NR activity. With this view the activity of NR was significantly (p≤0.05) varied according to different nutrient combinations (Figure 1).
|
|
.png)
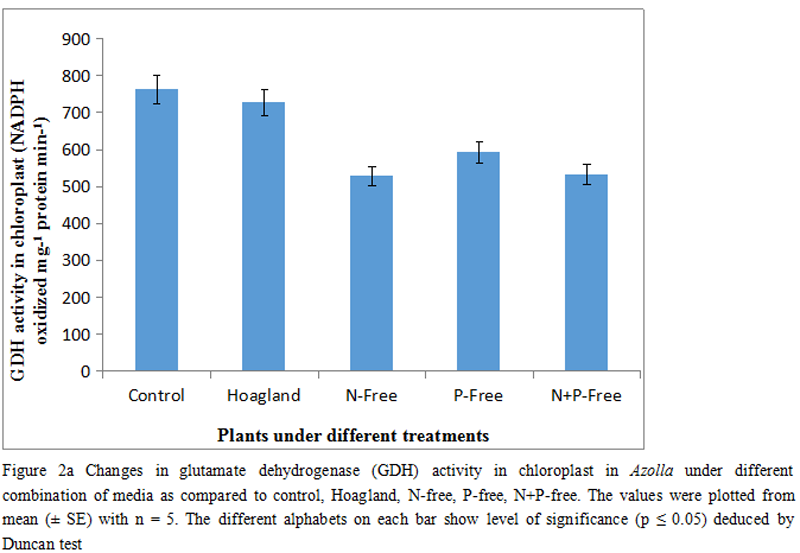
Plants responded well under artificial nutrients and recorded 1.09 fold in enzyme activity. Still, the other treatments including N, P, and both N+P free treatment had hardly shown any significant changes. The declines in activities were 6.25%, 3.38% and 8.51% respectively. Inorganic nitrogen when attains there lower redox (NO2), it showed that tendency to accept an H+ in the formation of NH4+. In plant system the activity of GDH with its two alternative forms (NADPH/NADH form) are attained more focus with NH4+ metabolisms (Figure 2a &2b).
|
|
|
|
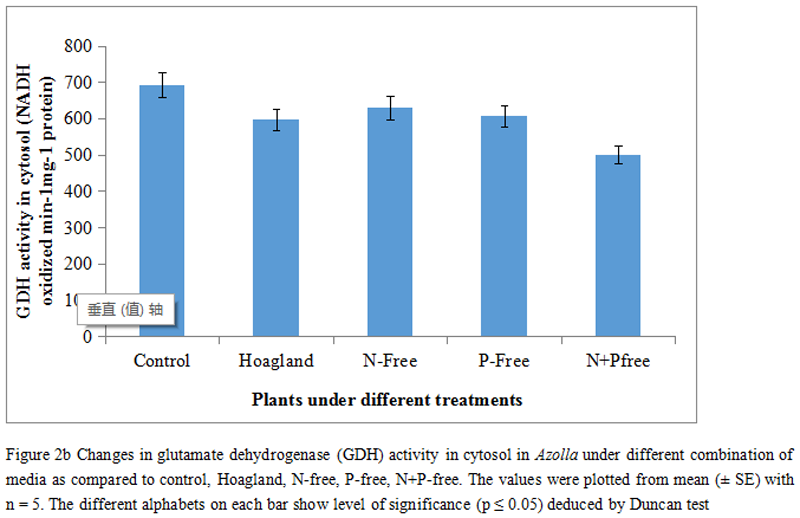
Thus, in the present experiment the chloroplastic form of GDH initially had shown a small decline, however, not significant (p≤0.05) by 4.73% and thereafter changed with 30.82%, 22.36% and 30.14% respectively (Figure 2a). As plants proceeded through other treatments, it recorded a significant down regulation of the activities. The minsimum activity was found with 30.82% fall as compare to control but the activities were moderately regulated, In comparison to control those were 22.36% and 30.14% respectively. Another isoform of GDH (NADH linked) were almost similar in trend and there by recorded a significant downhill in activities regardless of treatments (Figure 2b).
Plants were mostly suffered when both N and P were withdrawn as recorded with 27.69%. Moreover, the changes between N and P free set of nutrient were not statistically significant as compare to control by 9.003% and 12.46% respectively. In fact, plants under synthetic media had not improved in activities, rather decreased by 13.93% (Figure 2b). In another mode of nitrogen metabolism, Azolla plants were evaluated for inter conversion of aminso acids and organic acid pool by studying GOGAT activities (Figure 3).
|
|
.png)
Initially Azolla responded well in GOGAT assay with changes in absorbance at 340 nm. Interesting to note that the significant changes were most notable in the plants in complete synthetic media (Hoagland) as well as in the set withdrawn of N+P as compared to control. The changes were 1.26 and 1.39 fold respectively. On the other hand the changes were marginal, however, not significant (p≤0.05) under N and P depleted media and recorded 1.041 and 1.018 fold in accordance.
Glutaminse synthase (GS) becomes the crucial check point of nitrogen metabolism mostly transformation of aminso acids to its corresponding aminses. This enzymes, however, occurs in multi forms according to tissue specificity and reducing equivalent (NADH/NADPH). Still in present case the pool of GS was considered (Figure 4) a significant variation showed the plant's interactions differentially in sets of media amongst those. P-free and N+P-free were the most strikingly varied by 59.22% and 26.30% lesser than control. Moreover the up regulation of GS activity was maintained in maximum with 1.74 fold in case of Hoagland's media. On the contrary, plants grown in N-free nutrients had sustained the activity at par with the control. As a whole, with all the enzymes activation under various nutrients, plants were finally targeted to accumulate total protein. The activities of enzyme were not consistent in total protein accumulation. Thus, with this view Azolla were mostly suffered for protein accumulation al-through the treatments except complete media (Hoagland) (Figure 5). Therefore, the maximum amount of protein accumulated was 1.23 fold over control at complete media (Hoagland) and minsimum in N+P free media (34.48%). Other sets (like N and P free media) reduced the protein content with intermediate values of 1.29% and 11.4% respectively.
|
|
.png)
|
|
.png)
The differential behavior of the dry matter accumulation is reflected with the responses of plants to different nutrients availability and probably there with the assimilation in dry matter. Thus, in pure (de-ionized) water Azolla plants could not induce the growth over its existing biomass. It might sustain through maintenance of respiration like metabolic pathways. On the contrary, nutrient enrichment through Hoagland’s media was more promising for acquisition of dry matter over control. The dry matter accumulation under varying combinations of nutrient has already been documented in (Figure 6).
|
|
.png)
Now, on metabolic basis the dry matter accumulation is a resultant of carbohydrate metabolisms through acquisition of sugars left over utilization on catabolism (respiration). Initially Azolla plants were varied significantly (p≤0.05) when total carbohydrate was analyzed by anthrone method (Figure 7).
|
|
.png)
Plants accumulated well to continue their sugar accumulation all through the nutrient treatments. This got more expressed when the plants had to pass through complete Hoagland and N+P free media for 14 days. The variations showed in total carbohydrate that was significant at p≤0.05. On the contrary, plants under N and P free media were declined in carbohydrate status. The fall in carbohydrate, however, not significant and were 14.28% and 5.0% respectively. Plants responded well in current photosynthates like different component of reducing sugar under varying nutrient combinations. Thus, the pool of total reducing sugars recorded significant variantions (p≤0.05) among the treatments (Figure 8).
|
|
.png)
The maximum accumulations of reducing sugars were in the order of 1.83 and 1.63 folds respectively in complete (Hoagland media) and N+P free media. On the other hand, interestingly the depleted media (N and P free) were decreasing in trend. The decline in sugar content were 31.06% and 4.91% respectively in N-free and P free.
4 Discussion
From the facts and figures of the present experiment Azolla plants recorded some interesting features in their growth and metabolic activities as they interact among different combination of nutrients. It is quite in agreement that nutrients, particularly, NPK is integral minseral requirements primarily in assistance for basic life processes. Thus a significant variation has been in observation particularly in crop species for their impeded growth and development (El-Shahate et al., 2011). However, Azolla an aquatic pteridophytic species might be some exception to recover or sustain or even modify their cellular response according to stabilize the growth under such condition. Possibly, this may attribute for sustenance of Pteridophytic species even under more harsh conditions of habitat. As could be expected the deficiency of nitrogen or phosphate could make a disruption for normal chief cellular metabolic pathways. Both of those are played as the keys of growth and development. It is the efficiency of Azolla plants either in absence of nitrogen/phosphate could also maintain the basal level of growth. The nutrient disorder either of its non-availability (or even abundance) sometimes, however, for time being could resume the growth. It is the utilization of existing metabolites and thus compensates the growth (Forni et al., 2001). With this Azolla plants in the present experiment could be assumed to overwrite the growth against N and P-deficit media. Still, interesting to note that withdrawn of P had sensitized more than N and thus a prediction for nitrogen remobilization from the pool of metabolism is established. For the phosphate might not be realized in limiting the growth (Lefebvre et al., 1990). The variations those are elucidated from the present experiment in plant’s performances for nitrogen metabolizing enzymes were highly significant both at p ≤0.05 and p≤0.01. The nitrogen content in plant tissues as well as its direct or indirect reflection in different enzymatic activities, in fact, correspond the balance of photosynthetic reduced carbon and inorganic nitrogen-assimilation. Azolla in the present experiment is much ahead of nitrogen reserves than carbohydrate pool both of its own assimilation as well as from symbiotic partner. This over accumulation of total nitrogen mostly the aminso-nitrogen was higher among the plants where P-deficiency is lower. Thus, we found the nitrogen accumulation of Azolla species remained higher, probably to maintain a good biomass acquisition. This may be indicative of the fact that higher nitrogen gain would have been governed initially by the photosynthetic surplus. These situations become more prevalent in earlier results where P-deficit situation dominsated the soil types (Sadeghi et al., 2013). With this findings the present experiment is also supported with some significant changes in nitrogen metabolizing enzymes initially the plants were evaluated with NR activities and scored a significant changes while the assays were monitored from different treatments.
Thus, the peak activity under complete nutrient media is referred to be the full supplementation for normal metabolic paths including nitrate reduction. These also add the constitutive demands for aminso nitrogen through initial stages of nitrate reduction. Interestingly, the three other combinations (N-Free, P-Free and N+P-Free) had hardly shown any significant variations amongst those, however, reduced the activity as compare to control. It assumes that any of the essential nutrients (like N or P) might be limiting the nitrogen source and therefore the pool of total nitrogen is not allowed to be converted into aminso-nitrogen. This is more supportive for the fact that nitrate reductase (NR), an inducible enzyme (Rai and Rai, 2003). In downstream reactions for nitrogen conversion into organic moieties is the synthesis of aminso acids like glutamate etc. Plants exercise the incorporation of NH4+ into carbon skeleton (keto acid) is initially accomplished by two isoforms of GDH (Lindblad et al., 1987). In the present experiment the activity of GDH in-vitro being extracted and purified from chloroplast recorded a significant decline although the combinations of N/P supplementations. The insignificant changes of GDH (chloroplast type) under control and Hoagland media undoubtedly suggest the compatibility of the all ingredients of the media on activity of GDH. On the contrary, the other three media deficits with N, P and N+P had maximally curtail the GDH activity in proportionate manner. GDH irrespective of any forms (either cytosolic or chloroplastic) requires a adequate amount of electrons flow from the Photosystem-II (PSII) in the form of NADH/NAD(P)H. Plants deprived from any short of nutrients may be reflected primarily on energy mediated carbon reduction as well as the reaction paths for energy yielded metabolisms.
It assumes that any of the essential nutrients and their absence (like -N or -P as in the present case) might be limiting the nitrogen source and therefore the pool of total nitrogen is not allowed to be converted into aminso-nitrogen. Plants deprived from any short of nutrients may be coupled with impaired photosynthetic electron flow as well as photo phosphorylation (Ito and Watanabe, 1983). Thereby, the product of later may limit the releasing equivalents as well as ATP to pull the reaction in completion of NH4 incorporation in keto-acid. Metal stress which replaces the adequate nitrogen acquisition in plants from soil also establish a down regulation of GDH activity (Raja et al., 2012 ). In correction to keto-acid and aminso acid inter conversion it is the transaminsation reaction where frequent reversal reactions between aminso acid and keto-acid supports the plants demands for those two. The glutaminse-2oxoglutarate aminso transferase (GOGAT) is characterized in plants where abundance of ATP finds a path to be exercised in conversion of keto-acid from organic aminses (Arrora et al., 2003). Under depletion or special supplementation of any nutrients may relief the plants induced toxicity from over accumulation of aminses in the tissues. With this Azolla responded in a noticeable way to change the pattern of GOGAT activity under different sets of treatments. Thus, the falls in enzyme activity in-vitro under synthetic media of N-free and P-free were almost similar. This possibly indicate the absence of nitrogen or phosphate could access to retain more aminses (in the form of glutaminse) to compensate the organic nitrogen deficiency (Singh et al., 2008). However, for the P-deficit set it would be the inadequacy of ATP in carrying out the reactions. The activity of GOGAT was regained when plants were supplemented with additive doses of N+P along with normal nutrients. The over accumulation of aminse through glutaminse biosynthesis pathway must have been toxicated beyond the cellular pH and thus the activity of GOGAT was induced to release more NH3 and proportionate accumulation of the keto-acid. It is the plant's potential to set free more aminso acids from their aminsated forms and thereby facilitating the biosynthesis of protein in constitution of stress tolerance.
The glutaminse synthase is offered in plant system with reversible inter conversion between glutamic acid and its corresponding aminses, glutaminse (Parrado et al., 2008). It is the plant's demand to replenish the pool of aminso acids while excess aminses may face the toxicity beyond cellular threshold. In the present experiment plants starve with nitrogen, phosphorus and both N+P recorded a distinguishing trend as compared to control. In close view plants without P are mostly affected with GS activity. This might be attributed to the phosphate requirement either in the form of supplementation of ATP or NAD(P)H (as reducing equivalent) in cellular system. In another way plants without N+P could retrieve the GS activity over P-Free treatment. Assuminsg the requirement of nitrogen source for aminso acids to aminse inter conversion the cartel of GS activity by phosphate deficiency would be recovered. In the plants supplementation of more nitrogen had sustained the GS activity in full space as compared to control found in Azolla plants. This is more established as plants recorded an over expression of GS activity under synthetic media (Hoagland) supplemented with different nutrients (including N and P) in excess.
The carbohydrate profile and its different fractions are the reflectants of photosynthetic activities and its concomitant inter-conversion in different sugar moieties. Plants under different deficits of nutrients: N-free, P-free, N+P free had experienced; However, seeminsg differently with up regulation in carbohydrate content. This is more inconsistent in comparison to common phenomenon of stress induced inhibition of photosynthetic reserves. The nitrogen and phosphorus are two essential elements in metabolism could somehow down regulate the nitrogen metabolism, still, plants could divert more carbohydrate pools in synthesis and their turn over in different sugar residues. In earlier studies it has been reported that N and P deficit stress might lead some left-over accumulation of carbohydrates (Mahasneh et al., 1996). Azolla in the present experiment had sustained the total carbohydrates consistently. In ferns, somehow results in proliferation of the vegetative growth may take place through up regulation of carbohydrate content. Similar trend was also experienced in plants with regards to total reducing sugar in the present experiment. Understanding well that the reducing in a consequence of current photosynthetic rate per se or hydrolysed products of reserve carbohydrates, plants might be accessed in either way for stress tolerance. Nitrogen and phosphate deficit in soil lead, however, in few cases a sort of osmotic stress (Singh et al., 2010). Supplementation of reducing sugar, thus, might have some opportunities to be acted as osmoticum (Forn et al., 2008). In fact, whatever the status of carbohydrate either through photosynthetic surplus (excluding respiratory and photo-respiratory loss of carbon) or inter conversion of carbohydrate metabolic pathways. Finally, accumulations of dry matter are the result (Arrora et al., 2003). It is the total growth performance that scored by plants potential to restore acquisitions of dry matter as a whole is the consideration. Thus, as expected an up regulation of dry matter increase under synthetic media (Hoagland) is a satisfactory responses of plants under proportionate nutrient supplementation. On the other hand, the decline of the same under N, P and N+P free treatments are the logistic behavior for plants deficiency of such essential nutrients to affect the entire metabolic profiles. Thus, the impeded dry matter accumulation is a result.
5 Conclusion
The present work is represented with some preliminsary observation on a aquatic fern (Azolla pinnata R. Br.) with regards to nutrients variability. In fact this work signifies the possible physiological changes of pteridophytic species when encountered with N and P deficit. In brief if the nutrient deficiency is considered as stress imposition the fern species has the same ability to alter the nitrogen metabolism. This is evident from modulation in activities of different nitrogen metabolizing enzymes and their resultant in protein and aminso acid. On the other hand, this fern species, however, could sustain their carbohydrate metabolism. Still it affected in their dry mater acquisition. Taking together, it appears that both carbohydrate and nitrogen constituents for physiological processes restrict their growth under nutrient limitation by altering the nitrogen and carbohydrate metabolic paths in physiological processes.
Acknowledgement
This work is supported by the financial assistance from DST-PURSE programme, DST, Govt. of INDIA.
References
Arrora A., and Singh P.K., 2003, Comparison of biomass productivity and nitrogen fixing potential of Azolla spp, Biomass and Bioenergy, 24: 175-178
http://dx.doi.org/10.1016/S0961-9534(02)00133-2
Barik S.K., Sahu M., Sudha M., Purushothaman C.S., and Ayyappan S., 2000, Phosphatase producing microbes and phosphorus in aquatic ecosystems, Ind. J. Microbiol, 40: 83-102
Botrel A., Mange C., and Kaiser W.M., 1996, Nitrate reduction, nitrite reduction and ammonium assimilation in barley roots in response to anoxia, Plant Physiology Biochem, 34: 645-652
Bradford M.M., 1976, Anal Biochem, 72
El-Shahate R.M., El-Araby M.M.I., Eweda E.W., and El-Berashi M.N., 2011, Evaluation of the effect of three different pesticides on Azolla pinnata growth and NPK uptake, Journal of American Science, 7(1): 1020-1031
http://www.americanscience.org
Esposito S., Guerriero G., Vona V., Rigano V.M, Carfagna S., and Rigano C., 2005, Glutamate synthase activities and protein changes interaction to nitrogen nutrition in barley: the dependence on different plastidic glucose-6P dehydrogenase isoforms, J. Expt. Bot., 56: 55-64
Forni C., Chen J., Tancioni L., and Caiola M.G., 2001, Evaluation of the fern Azolla for growth, nitrogen and phosphorus removal from wastewater, Water Res, 35(6):1592-8
http://dx.doi.org/10.1016/S0043-1354(00)00396-1
Healey F.P., 1973, Characterization of phosphorus deficiency in Anabaena, J. Phycol, 9: 383-394
http://dx.doi.org/10.1111/j.1529-8817.1973.tb04111.x
Hedge J.E., and Hofreiter B.T., 1962, In: Carbohydrate Chemistry 17 (Eds Whastler L. And Be Miller J.N.) Academic Press New York
Hoagland D.R., and Arnon D.I., 1950, The water-culture method for growing plants without soil, California Agricultural Experiment Station Circular 347: 1-32
Ito O., and Watanabe I., 1983, The relationship between combined Nitrogen uptake and Nitrogen fixation in Azolla- Anabaena symbiosis, New Phytol, 95, 647-654
http://dx.doi.org/10.1111/j.1469-8137.1983.tb03528.x
Kar P.P., Mishra S., and Singh D.P., 2000, Variability in Azolla sporulation in response to phosphorus application, Biol. Fertil. Soils, 31 458
http://dx.doi.org/10.1007/s003740000276
Kwinta J., and Cal K., 2005, Effects of salinity stress on the activity of glutaminse synthase and glutaminse dehydrogenase in triticale seedlings, Polish J Envir Stud, 14: 125-130
Lefebvre D.D., Duff S.M.G., Fife C.A., Julien-Inalsingh C., and Plaxton W.C., 1990, Response to phosphate deprivation in Brassica nigra suspension cells, Plant Physiol, 93, 504-511
http://dx.doi.org/10.1104/pp.93.2.504
Loyla-Vargas V.M., and Jimenez E.S., 1984, Differential role ofglutamate dehydrogenase in nitrogen metabolism in maiz tissue, Plant Physiol, 76:536-540
http://dx.doi.org/10.1104/pp.76.2.536
Lindblad P., Rai A., and Bergman B., 1987, The Cycas rettoluta-Nostoc Symbiosis: Enzyme Activities of Nitrogen and Carbon Metabolism in the Cyanobiont, Journal of General Microbiology, 133
Mahasneh I.A., and Tiwari D.N., 1996, Phosphate uptake by the blue-green alga Anabaena M116 used as biofertilizer in deep-water rice, Cytobios, 86, 113-121
Miller G.L.,1972, Anal Chem, 31 426
http://dx.doi.org/10.1021/ac60147a030
Parrado J., Bautista J., Romero E.J., García-Martínez A.M., Friaza V. and Tejada M., 2008, Production of a carob enzymatic extract: Potential use as a biofertilizer, Bioresource Technology, 99(7): 2312-2318
http://dx.doi.org/10.1016/j.biortech.2007.05.029
Peters G.A., and Mayne B.C., 1974, The Azolla, Anabaena azollae relationship: 1. Initial characterization of the association, Plant Physiol, 53: 813-819
http://dx.doi.org/10.1104/pp.53.6.813
Peters G.A., and Meek J.C., 1989, The Azolla-Anabaena symbiosis: basic biology, Ann. Rev. Plant Physiol, Plant Mol. Biol., 40: 193-210
Rai A.K., and Rai V., 2003, Effect of NaCl on growth, nitrate uptake and reduction and nitrogenase activity of Azolla pinnata Anabaena azollae, Plant Science, 164: 61-69
Raja W., Rathaur P., John S.A., Ramteke P.W., and Dar R.A., 2012, Effect of monocrotophos on electrolytic leakage, proline content and nitrogen metabolism of floating Pteridophyte Azolla microphylla, The Holistic Approach to Environment, 2(3): 111-120
Sadeghi R., Zarkami R., Sabetraftar K., and Van Damme P., 2013, A review of some ecological factors affecting the growth of Azolla spp., Caspian J. Env. Sci., 11(1): 65-76
Singh S.S., Mishra A.K., and Upadhyay R.S., 2010, Potentiality of Azolla as a suitable P-biofertilizer under salinity through acid phosphatase activity, Ecological Engineering, 36(82): 1076-1082
http://dx.doi.org/10.1016/j.ecoleng.2010.04.022
Singh S.S., Singh S.K., and Mishra A.K., 2008, Na+ regulation by combined nitrogen in Azolla pinnata-Anabaena azollae symbiotic association during salt toxicity, Ecotoxicology and Environmental Safety, 69(1): 32-38
http://dx.doi.org/10.1016/j.ecoenv.2007.04.001
Singh Y.V., Singh B.V., Pabbi S., and Singh P.K., 2007, Impact of organic farminsg on yield and quality of basmati rice and soil properties, 3: 20-23
Vitousek P.M., Cassman K., Cleveland C., Crews T., Field C.B., Grimm N.B., Howarth R.W., Marino R., Martinelli L., Rastetter E.B., and Sprent J.I., 2002, Towards an ecological understanding of biological nitrogen fixation, Biogeochem, 57-58: 1-45
http://dx.doi.org/10.1023/A:1015798428743
Watanabe I., Berja N.S., and Del Rosario D.C., 1980, Growth of Azolla in paddy field as affected by phosphorus fertilizer, Soil Science and Plant Nutrition, 26(2): 301-307
http://dx.doi.org/10.1080/00380768.1980.10431212



. PDF(447KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. De A.K.
. Bera S.
. Adak M.A.
Related articles
. Nitrogen metabolism
. Carbohydrate profiles
. Nutrient deficiency
. Aquatic fern
Tools
. Email to a friend
. Post a comment